|
M. Ponce* y B. Trujillo* * Universidad Central de Venezuela Facultad de Agronomía.Departamento 1 trabajo financiado por el Consejo de Desarrollo Científico y Humanístico,Universidad Central de Venezuela. RECIBIDO: noviembre 14,1986. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
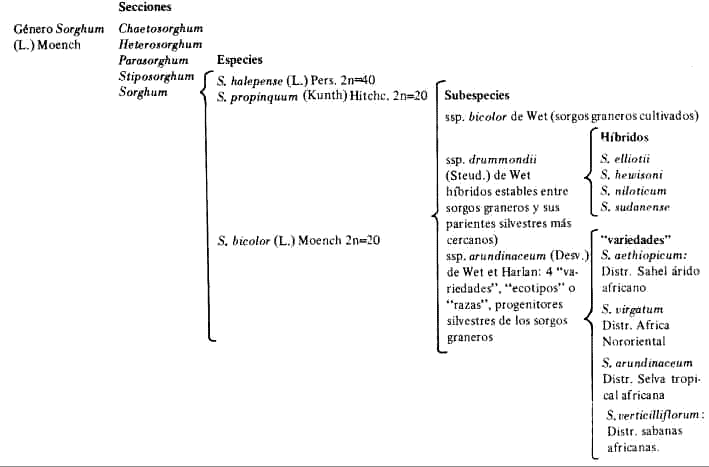
INTRODUCCIONA lo largo de la historia de la taxonomía del género Sorghum, se ha llegado a aceptar como funcional las secciones Eusorghum y Parasorghum de SNOWDEN (38, 39), pero no existe en cambio consenso entre los especialistas en cuanto a la clasificación de las diferentes entidades de la sección Eusorghum, la cual comprende al 'Falso Johnson'. Las diferentes posiciones involucradas pueden resumirse en dos extremes: una más antigua que consideraba todas las entidades de Sorghum que se iban describiendo como especies separadas, posición ésta que se mantuvo desde LINNAEUS (28), hasta STEUDEL (40), y otra integracionista que surge con ALEFELD (1) y es desarrollada por HACKEL (18). ALEFELD (1) ideó el sistema de subordinar todos los sorgos cultivados como variedades de una especie: Andropogon sorghum. HACKEL (18) amplió dicho sistema al incluir en el mismo los sorgos silvestres y propuso adoptar dos denominaciones subespecíficas: A. sorghum ssp halepense para las silvestres y A. Sorghum ssp sativas, para las cultivadas. También KOERNICKE (25) elaboró una clasificación semejante a la de HACKEL. Posteriormente se han realizado estudios monográficos del género limitados a la sección Eusorghum de SNOWDEN, o a subsecciones o series de la misma, las cuales interesan porque incluyen a la entidad 'Falso Johnson' y a otras muy afines a allá. Cabe citar a PIPER (34) quien estudió sobre todo a los sorgos cultivados en USA, tanto en poblaciones naturales como en material de herbario y extendió los estudios al material depositado en Kew y en Berlín. Como resultado de ello, adoptó una posición más o menos intermedia al considerar 11 subespecies de A. sorghum y ó subespecies de A. halepensis. STAPF (37) discutió las clasificaciones de HACKEL, KOERNlCKE y PIPER, enfatizando la necesidad de reconocer unidades más pequeñas y con ese criterio describió todas las entidades silvestres y cultivadas conocidas por él. Creó la sección Eusorghum Stapf donde incluye 27 especies, separando de éstas, por clave, a S. versicolor, S. purpureo-sericeum y S. dimidiatum, caracterizadas por canículas con ramas primarias simples, racimos terminales y vainas de los nudos superiores largo-barbadas. El trabajo taxonómico con criterio morfológico más complete, basado en material de herbario, es el de SNOWDEN (38, 39), quien adopta la denominación Eusorghum de Stapf pero pasa S. versicolor, S. purpureo-sericeum y S. dimiduatum a una sección aparte, Parasorghum, cuyas especies poseen un número cromosómico 2n = 10, a diferencia de las de la sección Eusorghum subsección Arundinacea que tienen 2n = 20 (KARPER y CHRISHOLM, 24; GARBER, 13). SNOWDEN contempla para la sección Eusorghum 31 especies cultivadas anuales (subsección Arundinacea, serie Saliva); 17 especies silvestres anuales y/o perennes (subsección Arundinacea, serie Spontanea) y 4 especies silvestres rizomatosas perennes (subsección Halepensia), lo cual hace un total de 54 entidades. Posteriormente DE WET et al. (5, 6, 7, 8, 9), en una serie de trabajos basados en los resultados y consideraciones de KARPER y CHRISHOLM (24), GARBER (13, 14, 15, 16), CELARIER (2, 3), DOGGET (10), LIANG y CASADY (27), replantean la monoespecificidad de los complejos cultivados y semisilvestres, al considerarlos como un solo conjunto con el nombre de Sorghum bicolor (L.) Moench y acoger, aparentemente, la clasificación de GARBER (14) y CELARIER (2) la cual subdivide el género Sorghum en cinco secciones: Chaetosorghum, Heterosorghum, Parasorghum y Stiposorghum, que comprenden las especies verdaderamente silvestres y Sorghum. En este último, DE WET contempla tres especies, dos perennes rizomatosas, de la subsección Halepensia Snowden y una anual, S. bicolor (L.) Moench, de la subsección Arundinacea Snowden, con taxones cultivados, silvestres y malezosos. De la serie Spontanea de Snowden, DE WET diferencia cuatro especies "snowdenianas" como híbridos estables entre los sorgos cultivados y miembros del complejo Spontanea: S. elliotii, S. hewisonii, S. niloticum y S. sudanense. El resto de los taxones "snowdenianos" los agrupa en las variedades" aethiopicum, arundinaceum, verticilliflorum y virgatum, para lo cual ha tenido que asimilar las faltantes a algunas de las "variedades" nombradas o a híbridos entre las mismas (Cuadro 1). Las nueve especies "snowdenianas" restantes se distribuirían entre las "variedades" anteriores, de la siguiente forma:
Además de los investigadores antes mencionados , han tenido que ver también con la taxonomía del complejo Sorghum, en relación con el problema planteado, KARPER y CHRISHOLM (24) quienes estudiaron el número cromosómico y realizaron experiencias de hibridación. Ellos sostienen la separación del género en al menos tres especies: S. halepensis, S. vulgare y S. versicolor, y agregan que se justifica la subdivisión de S. vulgare en varias subespecies cuando hay marcadas diferencias morfológicas. GARBER (14, 15, 16), subdividió a Sorghum en seis subgéneros, contemplando entre ellos a Eusorghum como correspondiente con la sección Eusorghum Snowden. El número cromosómico básico dentro del género Sorghum de GARBER es 5, el cual se encuentra en sus subgéneros Parasorghum y Stiposorghum, los cuales a su vez tienen miembros tetra y hexaploides. El número haploide de su Eusorghum es 10, con miembros diploides y tetraploides, y el número haploide de Chaetosorghum y Heterosorghum es 20. DOGGET (10, 11) considera a toda la sección Arundinacea Snowden como una especie compleja, modificándose por introgresión con los varias tipos cultivados y quizás con S. halepense, y agrega que por esa vía está sometida a un proceso de especiación o especiaciones. Mantiene que no hay argumento serio que impida reconocer que los sorgos silvestres y cultivados difieren a nivel específico, como lo demostrarían los estudios de índice de hibridación en Africa Oriental, donde se tiene una impresión básica de tres poblaciones distintas que dichos estudios corroboran y además que el cultivo de algunos híbridos naturales, que parecían ser verdaderos antecesores intermedios, mostraron esterilidad en F3, lo cual es indicación de alguna ruptura genética en el cruce. LIANG y CASADY (27), estudiaron 22 caracteres para 21 especies "snowdenianas" de la subsección Arundinacea, series Saliva y Spontanea, a fin de observer el o los patrones de variación morfológica y pudieron agruparlas en tres series, la primera correspondiendo con la serie Saliva Snowden, la segunda correspondiendo con la serie Spontanea Snowden y la tercera constituida solo por S. aethiopicum, no correspondiendo con ninguna de las anteriores, pero sí con características intermedias, por lo cual la consideran probablemente híbrida. Señalan que los datos no responden la pregunta de si las especies "snowdenianas" son especies, subespepecies o razas, respuesta' que debería descansar sobre estudios genéticos y citogenéticos más cuidadosos. Concluyen que quizás las entidades estudiadas sean "especies morfológicas" o grupos de individuos que se semejan entre sí en la mayoría de sus caracteres visibles, y que se debe estar consciente que las especies no son unidades estáticas sine estadios en el proceso de divergencia evolutiva, así como del hecho de que la interferencia del hombre con los habitats puede conducir concebiblemente a la eventual obliteración de la separación de las especies. MURTY et al. (33), estudiando la colección mundial de Eusorghum Snowden que contiene más de 10 00.0 entradas, manifiestan que no pudieron agruparlas de manera claramente definida, pero que no obstante, usando diversos tipos de análisis, llegaron a obtener nueve categorías principales a las cuales asignaron rango específico JAKUSHEVSKY (23), reduce a nueve las especies "snowdenianas" de Sorghum. En lo que atañe a la investigación taxonómica en Venezuela, aparecida en la literatura especializada, el primer señalamiento de un taxon, hay día ubicado en Sorghum, lo hace KUNTH (26) bajo el nombre de Andropogon decolorans, el cual ya había sido descrito como Holcus decolorans por WILLDENOW (41) y fue transferido a Sorghum por ROEMER y SCHULTES (36). Todos los nombres citados han pasado a la sinonimia. A este respecto, SNOWDEN (39) hizo una revisión de los mismos y señala que HACKEL (18) los ha incluido en su Andropogon sorghum var. effusus, el cual a su vez es sinónimo de S. arundinaceum (Desv.) Stapf, pero advierte que es evidente que HACKEL trató como A. Sorghum var. effusus a más de un taxon, entre ellos, S. macrochaeta, S. arundinaceum y probablemente S. verticilliflorum. ERNST (12) señaló la presencia de cultivos escasos de " sorgo escobero de panoja muy desparramada", los cuales incluye dentro de Sorghum vualgare. LUCES (29) hace mención de S. decolorans para señalar que no obstante ser tan común en Brasil, no se le ha vuelto a encontrar en Venezuela. En un trabajo sobre malezas PRIETO (35), proporciona una descripción e ilustración de la planta, con el nombre de Sorghum arundinaceum Stapf. Una revisión de las muestras actualmente existentes en los herbarios VEN. y MY. permite señalar que gran parte de las mismas se encuentran denominadas como Sorghum bicolor var. arundinaceum, acogiéndose a la terminología de DE WET. Cabe señalar que en la literatura científica no agrostológica como es la fitopatológica y malezológica, la planta aparece citada unas veces como S. arundinaceum y otras como S. verticilliflorum (MALAGUTI, 30, 31, 32). De lo expuesto se concluye la necesidad que existe de dilucidar tanto el rango como la denominación correcta de una entidad vegetal muy perjudicial, pues es infestante de campos de producción de semillas al cruzarse con la línea androestéril utilizada para tal fin Agregase a ello el ser hospedadora de plagas y patógenos que afectan grandemente al sorgo granífero en la medida en que se extiendan las áreas cultivadas, principalmente de sorgos graneros, arroz, caña de azúcar y pastos. MATERIALES Y METODOSSe utilizaron exsicatas y muestras vegetales frescas de 'Falso Johnson', provenientes de diferentes zonas del país y muestras de plantas cultivadas expresamente. Para su observación se usó lupa estereoscópica con lente Zoom de hasta 25x, glicerina, agudas de disección, etc. Para los cultivos se utilizó el área de Flora Biológica del Jardín Botánico Universitario Maracay, la cual dispone de canteros a cielo abierto que permiten el estudio en condiciones subóptimas semicontroladas. El material de herbario revisado proviene del MY., enriquecido con las muestras tomadas expresamente. Se añadió pocas muestras disponibles en VEN., ya que la mayoría de las mismas se encontraban en el exterior en calidad de préstamo. Se confrontó el material vegetal de herbario con la literatura pertinente y, en especial, con las obras monográficas de STAPF (37), SNOWDEN (39),, DE WET et al. (ó, 7), lo que resultó insuficiente para arribar a una conclusión. Además se examinó más de 150 ejemplares provenientes de poblaciones naturales y poblaciones cultivadas con la finalidad de aumentar la información sobre su distribución geográfica en el país, su rango habitacional, la posible plasticidad fenotípica asociada al rango habitacional y estabilidad morfológica de las poblaciones y visualizar posibles poblaciones o individuos híbridos de 'Falso Johnson', intentando detector al otro progenitor y grado de ocurrencia, fructificación y viabilidad de las posibles progenies de los híbridos. Para ello se procedió a la elaboración detallada de una descripción morfológico-agronómica, considerada representativo de la entidad 'Falso Johnson', así como al reintento de identificación de la entidad según clasificaciones de STAPF (37), SNOWDEN (39) y DE WET (6, 7). Para cubrir estos objetivos se procedió a aumentar el muestreo de la entidad en el país, mediante un registro de datos sistematizados que tomara en cuenta caracteres morfológicos "snowdenianos" y no "snowdenianos" y caracteres agroecológicos. Así mismo, se obtuvieron propágulos de individuos-promedio considerados representativos de las poblaciones típicas, con fines de siembra tanto en condiciones subóptimas de luz, agua, nutrimentos y sin competencia, como en condiciones de déficit hídrico. Por otra parte, se hicieron no sólo observaciones sistematizadas de campo de posibles híbridos y de su grado de ocurrencia, sine que éstos se cultivaron para observer la generación siguiente; igualmente, se procedió a probar la posible hibridación, bajo condiciones de cultivo, entre S. bicolor ssp. bicolor macho estéril ATX.399 y la entidad 'Falso Johnson', para observer la variabilidad morfológica en muestras de 30 individuos, así como también la viabilidad de los mismos. RESULTADOSLas observaciones sobre distribución permiten afirmar que el 'Falso Johnson' ha sido señalado en casi todas las entidades político-territoriales del país, exceptuando quizás el estado Nueva Esparta (HOYOS, 21) y las Dependencias Federales del Caribe (Islas). No obstante predominar en pisos bajos, ha sido colectado desde el nivel del mar hasta los 1 300 m, en Boconó, estado Trujillo. En cuanto a su distribución por tipos de vegetación, se han registrado enclaves en espinares degradados (Quíbor, estado Lara), pasando por selvas deciduas, semideciduas y en galería, hasta selvas macrotérmicas siempre verdes intervenidas (oeste del lago de Maracaibo, estado Zulia y San Carlos de Río Negro, Territorio Federal Amazonas), regularmente en lugares perturbados abiertos, ya como áreas cultivadas o sucesionales, y casi siempre sobre formas de relieve plano: planicies, terrazas fluviales, etc En referencia al rango habitacional, se comporta generalmente como heliófita, lo cual se traduce en la configuración del biotipo: macolla cilíndrico-obcónica o abierta con hojas verde-amarillas y en ángulo abierto, aun cuando puede llegar a comportarse como hemiesciófila; y como invasora, ya que en los últimos 23 años se ha extendido a casi todo el territorio nacional. Respecto a las condiciones hídricas, se le ha hallado en suelos desde condiciones de saturación hasta secos y excepcionalmente muy secos, siendo lo más frecuente encontrarla en condiciones subhúmedas a húmedas, de textura franco-arenosa hasta arcillosa, pero es más común observarla en suelos francos de alto fertilidad. La expresión morfológica es en general constante, si se exceptúan los extremos de insuficiencia lumínica, hídrica y nutricional que afectan al fenotipo disminuyendo el porte, el grado de macollamiento y/o el llenado de espiguillas; en el último case repercutiendo sobre la forma de la inflorescencia que aparece con raquis y ramas más bien rectas, semiaplicadas o aplicadas y no arqueadas o ascendenteflexuosas como es común. Por el contrario, en condiciones de cultivo con plena exposición solar, balance hídrico favorable, alto fertilidad y carencia de competencia, se llegó a observer porte de hasta 2,20-2,40 m, con inflorescencia de 40 x 35 cm y hasta 50 macollos basales. No llegó a observarse verdaderas poblaciones de híbridos entre 'Falso Johnson' y entidades afines. Esporádicamente se apreciaron individuos aislados, morfológicamente diferentes (híbridos naturales F1?), generalmente cercanos a sitios de cultivo de sorgos graneros y cuyas características se presentan resumidas en el Cuadro 2. Los caracteres exhibidos por los posibles híbridos F, permiten conjeturar que en los mismos están presentes tanto el vigor híbrido (porte, dimensiones de panículas, longitud de láminas, etc.), como posibles caracteres intermedios entre 'Falso Johnson' y Sorghum bicolor ssp. bicolor (densidad de macollamiento, anchura de láminas, densidad de panículas y con ésta el número de nudos por inflorescencia y por verticilio, además, ancho de espiguillas, caducidad de las mismas, etc.). Son también visibles caracteres más próximos a uno u otro posible progenitor; de cultivares graneros se observe el grosor de tallos, color y ancho de hojas, raquis erecto, raquis y ramas de la inflorescencia rígidos, verticilación o subverticilación de las ramas de la panicula y del 'Falso Johnson', longitud de espiguillas sésiles y parcialmente forma, longitud de las glumas y consiguiente superposición de las mismas hasta el ápice. Es necesario resaltar la muy baja proporción de espiguillas fértiles encontradas, lo cual explicaría en parte que sea casi excepcional hallar descendientes de éstos en condiciones de campo. La progenie obtenida del híbrido anterior (posible F2 ) en condiciones de cultivo resultó ser heterogénea (Cuadro 3), aun cuando la mayoría de los individuos fueron similares en conjunto, a primera vista, al 'Falso Johnson'. La semejanza al 'Falso Johnson' se debió primordialmente al porte, forma de la macolla cilindroobcónica con ramificaciones en los nudos superiores no enraizados, entre 2-5 por individuo, alto densidad de macollamiento y alga de la forma de la inflorescencia (raquis a veces alga flexuoso y ramificaciones de la inflorescencia flexuosas) Un examen más atento, sin embargo, revela diferencias importantes en cuanto al diámetro del tallo, dimensiones y color de hojas, longitud de raquis y de ramificaciones de la inflorescencia, número de nudos/inflorescencia, tipo de ramificación de la inflorescencia, número de ramas/verticilo Es importante señalar que no se encontraron espiguillas llenas debido a la atrofia o crecimiento rudimentario del androceo y/o gineceo y más aún, a que en algunas inflorescencias las espiguillas no alcanzaban el crecimiento normal.
La posibilidad de cruce entre 'Falso Johnson' y S. bicolor ssp. bicolor se comprobó con la progenie homogénea obtenida a partir del cruce 'Falso Johnson' x cultivar macho estéril ATX.399 (Cuadro 4). Los resultados muestran un escaso vigor híbrido expresado en un porte y longitud de láminas ligeramente superior a ambos padres. Como caracteres intermedios se encontró: densidad de macollamiento, diámetro de tallos, anchura de láminas, grosor del raquis y ramas de la inflorescencia, densidad, longitud y número de ramas por verticilo de la inflorescencia; como caracteres próximos al macho estéril: forma de la macolla, número de nudos/inflorescencia, verticilación y subverticilación, ¿espiguillas más o menos persistentes?. Como caracteres próximos al 'Falso Johnson': color de las láminas, dimensiones de la panícula, dimensiones de la espiguilla (aun cuando la anchura es ligeramente mayor), superposición de las glumas hasta el ápice. En este híbrido no se encontraron espiguillas llenas observándo se , en general, aborto de dos anteras, la única presente inclusa y vacía; el gineceo por lo general normal pero no fecundado, y a veces ovario y estigmas rudimentarios o atrofiados. Lo anterior demuestra que el híbrido es estéril, factible de propagarse sólo por estacas. DESCRIPCIONPlanta anual, desde 412 meses según las condiciones de crecimiento. Porte desde 0,80-2,20 (2,40) m. Los individuos más frecuentes (malezas arvenses o marginales a los cultivos) alcanzan 1,20-1,80 m. Macollamiento monocaule (raro) por extreme deficiencia lumínica, a multicaule con hasta 20 macollos basales en condiciones dásicas, pudiendo llegar a 30-50 en condiciones de cultivo. Diámetro basal, 3-6 (- 10) cm, forma geniculado-ascendente en condiciones muy favorables de luz, en general cilindro-obcónica hasta erecta y cilíndrica en condiciones de alto competencia. Sistema radical subterráneo abundante, profundo y de raíces gruesas. Raíces fúlcreas presentes, pudiendo llegar a ser absorbentes. Tallos, 5-7 mm de (, normalmente ramificados. Hojas del macollo principal de 11-16, normalmente 4-6 (-8). Vainas desde 10-22 x 2 cm; las inferiores sobrepasando en longitud al entrenudo respectivo y las superiores sobrepasadas por los entrenudos; vainas prepedunculares y pedunculares muy grandes, las últimas 36-45 cm de largo; pilosas en la vecindad de la lígula, de color verde-claro excepto en los extremes, blanco-perladas internamente. Lígulas membranosas ancho-corto-triangulares, 2 mm de ancho máximo, microciliadas externamente. Láminas fasciarias sensu stricto (largo: ancho = 12 ó más), con dimensiones de 30-50(-60)x 2-3(-3,2) cm, en general de color verde-amarillento y excepcionalmente alga oscuro en condiciones de alto fertilidad y/o semisombra. Base breviauriculada y equilátera, vena media blanca de 2 mm o menos; posición con respecto al eje, variable con ]a edad: agudas cuando adultas y obtusas en fase senescente; pulimento opaco, indumento desnudo, pubescencia glabra, nervación principal alga prominente por el haz, con más o menos 16-18 nervios; mancheo por enfermedades, escaso o excepcionalmente abundante. Inflorescencias emergentes dispuestas en panículas laxas a muy laxas, conformando pirámides inequilateras de ápice incurvo, la cual varía con la edad; cuan
do muy juveniles el requis es erecto y ;as ramas emergidas alga flexuosas; cuando adultas el raquis es flexuoso y las ramas péndulas; cuando maduras y senescentes se observe el raquis irguiéndose progresivamente hasta llegar a ser erecto; dimensiones 37-40 (-50) cm de largo x 35 cm de diametro basal; pedúnculo cilíndrico, raquis ligeramente acanalado, delgado, 1,5-2,8 mm de diámetro, con 10-11 nudos; tipo de ramificación muy variable, basalmente verticilada, subverticilada, opuesta o alterna y suprabasalmente alterna u opuesta; las ramas más largas corresponden por lo general a las basales, alcanzando hasta 24 cm, ramificadas o no desde la base hasta unos 10 cm de la misma. Posición de las ramas basales, semiaplicadas y arqueadas y las superiores agudo-ascendente-oblicuas o ascendente-flexuosas; los racimos en orden tercero de ramificación, 2-6 por rama, comúnmente 45; 1,5-4,8 cm de longitud con 2-6 espiguillas; longitud de entrenudos, 4 mm Espiguillas sésiles articuladas, caedizas, callo cano-pubescente, pelos 1 mm longitud; de forma estrecho-eliptico-lanceoladas a ovado-eliptico-lanceoliadas, en general abultadas abajo y levemente deprimidas hacia arriba, llevadas en intervalos de 4 mm, 6,5-8,0 x 1,8-2,0 mm, generalmente 7 x 2 mm. Gluma inferior 7 x 2 mm, eliptico.lanceolada, biaquillada; dorso ligeramente redondeado en su mitad inferior, 9-11 nervada, los nervios mas conspicuos anastomosándose hacia el ápice; de consistencia crustácea en los dos tercios inferiores y papirácea en el tercio superior, ápice agudo a muy agudo, aparentemente acuminado por prolongación de las márgenes membranosas de las quillas, pulimento esplendente y pubescencia cano a rojizo-largo-hirsuta en los tercios media e inferior y glabra en el tercio superior; el viente con márgenes membranáceas espinuloso-cilioladas a lo largo de las quillas, 2-4 nervios; color generalmente amarillo en los dos tercios inferiores y blanco interrumpido de verde a lo largo de ;as venas en el tercio superior. Gluma superior ligerisimamente más corta y alga más estrecha, en general 6,9 x 1,8 mm, con el dorso más prominente, elíptico-lanceolado-aguda, naviculada, monoaquillada en el ápice, de consistencia crustácea con ápice papiráceo, de pulimento esplendente y pubescencia glabra excepto en el dorso donde es escasamente cano-pilosa, 3-5 nervía en el dorso y 2 nervia en el vientre, el cual es de márgenes membranáceas; con color castaño claro y ápice blancuzco aunque verde en las venas. Lemma del flósculo inferior desde 5-6 x 1-1,7 mm semilageniforme (en un plano, elíptica, atenuada hacia el ápice) en donde casi la mitad inferior es semiventruda y la mitad superior es isósceles triangular, de modo que si se extiende en la mitad inferior alcanza hasta 2,2 mm de ancho, ápice ligeramente bífido, márgenes involutas, consistencia hialina, pubescencia cano-largo-ciliada. Pálea ausente. Flósculo superior con lemma hialina desde 3,3-4 x 1~1,3 mm, comúnmente 3,5 x 1,2 mm semilageniforme en un plano, ovada abajo luego elíptica, y profundamente bifida superiormente, alcanzando 1,5 mm de ancho, con lóbulos de 0,9-1,2 mm monoaquillada, ciliada, cano-Fargo-hirsuta en los lóbulos y el resto glabra, finamente nervada, aristada o no incluso en una misma inflorescencia; ;a arista es un haz de venas, prolongación del cuerpo central que excede o no los lóbulos, cuando corta es recta y si alga largo, retorcida y geniculada, castaña hacia la base y más ciara hacia el ápice, alcanzando desde 1-9 mm, caduca Pálea hialina de 3-3,8 x 0,5-0,8 mm, subtriangular-lanceolada, ancho-truncada en el ápice, largo-hirsuto-ciliada en el tercio superior, finamente 7-venada. Lodículas rodean al ovario lateralmente aproximándose por la cara ventral y manteniéndose distantes por la cara dorsal, ligeramente obtrapezoidales, subigualmente bicornutas en los extremos superiores, el cuerno interno más largo opuesto a la cara ventral, con base de 0,4 mm, ápice de 0,7-0,8 mm y altura de 0,6-0,7 mm; consistencia alga crasa y transparentes cuando jóvenes, alga coriáceas y castaño-oscuro cuando senescentes; de pubescencia glabra aún cuando el cuerno interno es largohirsuto. Anteras subdorsifijas, versátiles; de 2,5-3 x 0,8-0,9 x 0,3-0,4 mm, color variable con la edad, cuando adultas castaño claro en la base y amarillo-verdosas hacia el ápice, con tecas ligeramente desiguales, prismático-obtruladas, dehiscentes por hendidura apical. Estilos de 2-2,5 mm, estigmas desde 1,5-2,5 x 2-2,8 x 0,6 mm, plumosos y amarillos cuando adultos y rojizos cuando senescentes. Ovario de 1,2-0,8 mm. Grado apreciable de cleistogamia; los estigmas emergen primero, pero cuando ello ocurre ya ha habido fecundación y consiguiente inicio de formación del fruto, cariópside obovoide-eliptico, con cara dorsal alga aplanada, y cara ventral convexa alga rebordeada en el ápice, midiendo 2,7-3 (-3,4) x 1,41,9 (-2,0) x 1 mm, color castaño-claro. Marca embrollan subepidérmica, de contorno oblongo-elíptico, de 1,3-2 x 0,7-1 mm con embrión elipsoidal visible por transparencia. Hilo basal puntiforme, ligeramente estrecho inferiormente. Espiguillas pediceladas únicas o en pares cuando acompañan a la espiguilla apical de los racimos; son articuladas, prontamente caedizas, generaImente con pedicelos aplanados de 4,5 mm denso-largo-ciliados, canos o rojizos de callo largo-hirsuto; elípticoagudas, de 5-8,5 x 0,7-1,2 mm, ápice agudo, amarillo-verdosas. Gluma inferior biaquillada, papirácea a lo largo de los nervios y membranácea entre ellos, la membrana central prolongándose más allá del ápice, con márgenes involutos y escariosos en las quillas, 7-9 nervada. Gluma superior muy corta (2,5-4 mm) 4-ó,8 x 0,9-1 mm, frecuentemente excedida en 2 mm por la gluma inferior; estrecho-elíptico-lanceolada, navicular, monoaquillada, membranácea con márgenes involutos escarioso-ciliados espinulósula a lo largo de las quillas, 5-7 nervada. Flósculo inferior neutro o raramente masculino, generalmente con lemma ausente y cuando presente es estrecho elíptico-lanceolada, hasta 5,2 x 0,6 mm, hialina. Pálea ausente. Lodiculas en general ausentes y si presentes, rudimentarias. Flósculo superior neutro, masculino o raro morfológicamente hermafrodita con gineceo rudimentario. Lemma estrecho-elíptico-lanceolada de ápice agudo, desde 3,84 x 0,4-0,8 mm, bisquillada, escaso. Largo-hirsuto-ciliada en márgenes de las quillas. Lodículas presentes. Pálea ausente o si está presente, hialina, escasísimamente largo-hirsuto-ciliada. Estambres ausentes o presentes solo los filamentos y cuando hay anteras éstas son poliníferas de 2,5-0,35 mm. Gineceo ausente o si presente, rudimentario. REINTENTO DE CLASIFICACION DE LA ENTIDAD "FALSO JOHNSON' SEGUN CLASIFICACIONES DE STAPF (37), SNOWDEN (39) Y DE WET (6, 7, 9).Se confrontaron los caracteres morfológicos del material estudiado con los de la clave y descripciones de STAPF (37) (Cuadro 5). Los resultados muestran que la entidad en estudio presenta grandes semejanzas con S verticilliflorum (Steud.) Stapf en porte, tamaño de hoja, raquis arqueado de la inflorescencia, forma del cariópside, etc. y comparte algunos caracteres con S, arundinaceum Stapf, tales como: laxitud de la inflorescencia, características de la arista, y parcialmente, longitud de la espiguilla sésil. Es de hacer notar que STAPF (37) reseña grandes afinidades entre S. verticilliflorumm y S. arundinaceum a los cuales separa por forma y tamaño de las espiguillas sésiles y por la presencia constante o no de la arista y sus características. El Cuadro 6 muestra una comparación de caracteres morfológicos del material estudiado según la caracterización de SNOWDEN (39) con especies "snowdenianas" afines: S. arundinaceum, S. vogelianum, S. brevicarinatum, S. Castaneum, S. macrochaeta y S. usambarense con el objeto de encontrar el grado de similitud. Allí, el símbolo (+) significa complete semejanza, considerada como superposición de rangos de caracteres cuantitativos en más del 80% o plena similitud con un carácter cualitativo. Hay que señalar que estos porcentajes están sobrestimados respecto al índice de Sorensen. El símbolo (+) significa desde 50% de superposición de intervalos hasta más o menos 80%. Los porcentajes de similitud aparecen en el Cuadro 7. En el mismo puede observarse el grado de similitud (sensu lato) respecto a los caracteres contemplados, en el orden siguiente: S. verticilliflorum> S. brevicarinatum> S. arundinaceum> S. usambarense> S. castaneum> S, macrochaeta> S. vogelianum>. No obstante la gran afinidad con S. verticilliflorum hay desemejanza de caracteres con el mismo en cuanto a la forma de la gluma superior y presencia y características de la arista También hay que resaltar como desemejanza algunos caracteres de intervalo estrecho cuya superposición bordea la discontinuidad, tales como anchura de láminas y dimensiones de espiguillas. Si se utiliza el mismo procedimiento con la finalidad de hallar el grado de similitud considerando la clasificación de DE WET et al. (6, 9) como simple reagrupación de las especies "snowdenianas", S. verticilliflorum, S. usambarense, S. castaneum, S. macrochaeta, S. brevicarinatum, es una "variedad" que denominan "verticilliflorum", se encuentra una semejanza de 93% entre ésta y el 'Falso Johnson' y al hacerlo con la "variedad' arundinaceum, que comprende las especies "snowdenianas" S. arundinaceum y S. vogelianum, se encuentra una semejanza del 53% con el mismo. No obstante esta reagrupación, es necesario señalar que se mantienen casi las mismas desemejanzas del 'Falso Johnson' con el S. verticilliflorum (Steud.) Stapf descrito por SNOWDEN, es decir, presencia o no de la arista y dimensiones de la misma y forma y dimensiones de la espiguilla sésil.
Al comparar el material estudiado con las descripciones de DE WET et al. (6) (Cuadro 8) el mismo podría tener semejanza con la "variedad" verticilliflorum en cuanto a dimensiones de láminas y panículas muy laxas, no así con el tamaño de la arista que seria similar a la "variedad" arundinaceum. No dan datos de las espiguillas sésiles. Igualmente podría tener semejanza con la "variedad" verticilliflorum en lo que respecta a las ramas de la inflorescencia poco numerosas, posibilidad de división de las mismas a lo largo de toda su longitud y tipo de habitat (DE WET, 1970). Parcialmente se asemeja en porte, longitud de hojas, pubescencia de las espiguillas sésiles, tamaño de inflorescencia Con la "variedad" arundinaceum, las semejanzas serían: ramas de h inflorescencia largas y flexuosas, longitud de la arista y, parcialmente, longitud de las espiguillas sésiles. Difiere de h variedad 'verticilliflorum" por h duración del ciclo de vida, anchura de hojas, raquis y ramas flexuosas desde jóvenes, arista sólo ocasionalmente presente y corta. Difiere a su vez de la "variedad" arundinaceum por la duración del ciclo de vida, porte, longitud y anchura de láminas, longitud de panículas y distancia de división de las ramas de la inflorescencia. Difiere de ambas, además de lo señalado para coda una, en el ancho y forma de las espiguillas sésiles EL PROBLEMA DE Andropogon decolorans Kunth. En cuanto a saber si el material en estudio ha sido colectado anteriormente en el país y/o confundido con otra entidad afin, es necesario hacer referencia a Andropogon decolorans Kunth, el cual ha sido sinonimizado con S. arundinaceum (Desv.) Stapf y con S. halepense (L.) Pers. Una revisión de la sinonimia más complete, STAPF (37) y SNOWDEN (39), así como la revisión de índices (Index Kewensis, etc.) conduce a Andropon decolorans Kunth =sorghum halepense (L.) Pers., sin embargo, SNOWDEN (39) llega a afirmar que la sinonimia de A. sorghum var. effusus Hack con Sorghum arundinaceum (Desv.) Stapf es sólo parcial y agrega que todo el material de Africa Oriental y del Sur, identificado así por HACKEL, corresponde a Sorghum verticilliflorum (Steud.) Stapf. Por otra parte, al confrontar las descripciones de Andropogon de colorans Kunth con Sorghum halepense (L.) Pers., sensu stricto (SNOWDEN, 39), en aquellos caracteres que lo permitían, se encontró semejanzas en 14 de un total de 15 de ellos. El carácter diferencial fue el de glumas ovadas en A. decolorans contra elípticas a subelipticas en S. halepense, Al respecto, es importante considerar un comentario de KUNTH (26) al final de la descripción original de A. decolorans que dice textualmente" . . Andropogon avenaceus o afin; difiere sólo por h ausencia de arista; o antes bien, ¿caducas en nuestra planta?", y en la página anterior, donde describen Andropogon avenaceus Schrad., se le asigna a éste como sinónimo el de Holeus halepensis,, que a su vez es sinónimo de S. halepense (L.) Pers. Agréguese que la descripción de A. decolorans Kunth, como las de la mayoría de la época, es imprecisa e insuficiente en caracteres y se basó en uno o quizás escasos individuos y no en poblaciones, en perjuicio de la expresión satisfactoria del intervalo de los caracteres, de modo que se puede considerar con un alto grado de factibilidad el paso definitiva de A. decolorans Kunth a la sinonimia de S. halepense (L.) Pers., incluso sensu stricto. No obstante, una seguridad satisfactoria amerita el examen del material tipo.
DISCUSION Y CONCLUSIONESSe evidencia en la revisión de literatura la gran disparidad de criterios que hay respecto al concepto de "especie" y al status taxonómico de las entidades integrantes del género Sorghum. Además, el 'Falso Johnson' ha sido identificado o le han sido aplicados inconsistentemente binomios correspondientes a diferentes jerarquias taxonómicas. Por otra parte, los resultados de la confrontación de caracteres del 'Falso Johnson' con las descripciones o caracterizaciones de las entidades consideradas, no permiten su ubicación satisfactoria en ningunas de ellas, ya sea por insuficiencia de información de las descripciones o porque efectivamente no concuerda con tales entidades. En vista de lo expuesto, parece conveniente discutir la aplicabilidad de los criterios involucrados en las clasificaciones, la bondad de las descripciones y la semejanza o desemjanza del 'Falso Johnson' con las entidades cuyas denominaciones ha recibido. Respecto a criterio de clasificaciones, hasta Stapf y Snowden, están funda mentadas en caracteres morfológicos externos, inicialmente sólo tipológicos y más adelante alga poblacionales, en donde, sin embargo, persisten las fallas inherentes a este tipo de clasificación: número de caracteres insuficientes y no siempre acompañados de su amplitud de variación cuantitativa, incidiendo ello en la dificultad de captar la variabilidad de la expresión fenotípica, en la inconsistencia en el uso de los mismos, lo cual restringe la comparabilidad en las confrontaciones y en la apreciación errónea en cuanto a la importancia de la presencia y/o manifestación de un carácter, por ejemplo, presencia de arista, distancia de ramificación de las ramas basales, etc. Además, tal criterio, sólo morfológico, es en sí insatisfactorio porque se limita únicamente a un aspecto de la biología de la planta. Algunos autores conceptualizan "especie" basados solamente en el libre flujo de genes o carencia de barrera genética, lo cual resulta insuficiente sobre todo al aplicarlo limitadamente a algunas pruebas de interfertilidad, no proseguidas en número suficiente de progenie, y desligada de otros aspectos de la biología y ecología de las plantas. En ese sentido HARLAN y DE WET (19) al caracterizar el "Conjunto Genético Primario", categoría informal que hacen más o menos equivalente a h especifica, señalan que ~ .... éste corresponde con el concepto tradicional de especie biológica. Entre las formas de este conjunto genético el cruce es fácil, los híbridos son en general fértiles, con buen apareamiento cromosómico, la segregación de genes es aproximadamente normal y la transferencia genética es generalmente fácil. La especie biológica incluye casi siempre razas espontáneas (silvestres y/o malezosas) así como razas cultivadas. Para aclarar esto, proponemos que la especie se divide en dos subespecies: subespecie A con razas cultivadas y subespecie B con razas espontáneas ". De acuerdo a los resultados obtenidos, no sería posible ubicar al 'Falso Johnson' en este "Conjunto Genético Primario", ya que el cruce natural no es tan fácil con los sorgos cultivados probados y los híbridos tienden a ser estériles. En cuanto al problema de asimilar o no las silvestres simpátricas afines a las cultivadas domesticadas, DOGGET (10) señala ".... por consiguiente podemos concluir que los sorgos cultivados fueron desarrollados a partir de los sorgos silvestres de Africa mediante selección disruptiva . . . " y más adelante: " . Los sorgos cultivados llegaron a ser distintos a nivel especifico y llegó a establecerse una población intermedia, separada, que ocupó cultivos abandonados y campos marginales" Al respecto, DAVIS y HEYWOOD (1963) señalan los uses varias del término especie, indicando que es empleado corrientemente en dos muy diferentes sentidos o definiciones para grupos de individuos que poseen atributos en común, principalmente morfológicos, y para poblaciones mendelianas o conjuntos genéticos reproductivamente aislados. Agregan que han dada también razones de por qué las especies deberían ser retenidas en su sentido tradicional como grupos de individuos reconocible morfológicamente, mientras que las especies de los conjuntos genéticos deberían ser denotadas por alguna otra terminología. Habría que agregar que en la conceptualización de especie es necesario tomar en cuenta su condición dinámica, no obstante el aislamiento genético, y además de éste, la distintividad morfológica perceptible en la discontinuidad y una singular concurrencia de caracteres. Con respecto a la posibilidad de asimilar las entidades silvestres y malezosas a las cultivadas domesticadas, es importante señalar que estas últimas no tienen vigencia por sí mismas en la naturaleza, ya que no soportan la selección natural, mientras que las malezas y silvestres afines, que persisten aún recibiendo introgresión genética, sí tienen vigencia como tales. En cuanto a los intentos de caracterización de las entidades, las de DE WET et al. no pueden considerarse verdaderas descripciones, ya que los datos morfológicos son muy escasos y presentados en forma poco ortodoxa, de modo tal que en este estudio se ha recurrido además a la sumatoria de las unidades "snowdenianas" que ellos proponen integrar, y con todo no se ha arribado a una identidad taxonómica satisfactoria, así que tomando en cuenta lo expuesto y basándose en los resultados obtenidos, se propone considerar las siguientes conclusiones: 1. Con criterio morfológico poblacional, correspondería al 'Falso Johnson' un rango taxonómico probablemente especifico, ya que al estudiar los caracteres de numerosas muestras de herbario y de muchos individuos de poblaciones vivas en: cultivos, áreas marginales y sucesionales, se ha encontrado siempre la persistencia de las poblaciones con la singularidad de caracteres que le confieren distintividad morfológica como aparece en la descripción que se acompaña, 2. No puede equipararse ni asimilarse su status taxonómico al de los sorgos cultivados, al menos los probados en este estudio, ya que éstos últimos ni tienen vigencia por si mismos en la naturaleza, ya que no soportan la selección natural, ni sostienen libre flujo genético con el 'Falso Johnson' 3. Tomando en cuenta la conclusión anterior, no sería correcto aplicarle la denominación de Sorghum bicolor subespecie arundinaceum. 4. Respecto a su ubicación taxonómica definitiva habría que contemplar: -la hasta ahora insatisfactoria identidad con los taxones afines; -el alto grado de afinidad mostrado con Sorghum verticilliflorum (Steud ) Stapf.; -el apreciable grado de afinidad mostrado con Sorghum arundinaceum (Desv.) Stapf y, en consecuencia, la posibilidad que corresponda con: a. Sorghum verticilliflorum (Steud.) Stapf, lo cual se comprobaría probablemente con el aumento de la representatividad de la expresión morfológica del mismo; b. híbrido estable entre S. verticilliflorum x S. arundinaceum, sujeto a comprobación mediante cruces de los mismos y estudios citogenéticos; c. alguna categoría subespecifica de S, verticilliflorum en proceso de especiación, 5. La ubicación definitiva está sujeta a estudios complementarios de la entidad, con criterio integral (biosistemático o biológico propiamente dicho), tomando en cuenta morfología, anatomía, palinología, citogenética, genética, genecologia, bioquimica, etc. AGRADECIMIENTOSe agradece al Ing. Agr. Hector Mena T. por sus valiosas sugerencias y el suministro del material macho estéril, así como también por la lectura del manuscrito . Al Prof. Víctor M. Badillo, por su ayuda en la citación bibliográfica y su atenta revisión del manuscrito. RESUMENSUMMARYBIBLIOGRAFIAALEFELD, F. Landwirthschaftliche flora oder die nutzbaren kultivirten garten - und feldgewachse mitteleuropa's . . . etc. 1866. n.v. fide House, l A. Guide to Sorghum breeding. ICRISAT, India. p. 6. 1980. CELARIER, R P. Cytotaxonomy of the Andropogoneae. III. Subtribe Sorgheae genus Sorghum. Cytología (Tokio) 23(4):395-417. 1958. CELARIER, R.P. Cytotaxonomy of the Andropogoneae. IV. Subtribe Sorgheae. Cytologia (Tokio) 24(3):285-303.1959. DAVID, P.H. y V.H. HEYWOOD. Principles of Angiosperm Taxonomy. New York. D. Van Nostrand Comp. 556 p 1963. DE WET, J.M.J. The origin of Sorghum bicolor, I. Morphological variation Bot. Gaz? 1967-a n.v. fide De Wet, J.M.J., J.R. Harlan y E.G. Price. Am. J. Bot.57(6):704-707.1970. DE WET, J.M.J. y J.P. HUCKABAY. The origin of Sorghum bicolor. II. Distribution and domestication. Evolution 21 :787-802.1967. DE WET, J.M.J., J.R. HARLAN y E.G. PRICE. Origin of variability in the spontanea complex of Sorghum bicolor. Am. J. Bot. 57(6 :704707. 1970. DE WET, J.M.J. y J.R. HARLAN. The origin and domestication of Sorghum bicolor. Econ. Bot. 25(2):128-135.1971. DE WET, J.M.J. Systematics and evolution of Sorghum Sect. Sorghum (Gramineae). Amer. J. Bot. 65(4):477-484.1978. DOGGET, H The development of the cultivated sorghum. In: Hutchinson, J.B. ed. Essays on crop plant evolution. Cambridge, Univ. Press. p. 50-59. 1965. 11.DOGGET, H. Sorghum. London. Longmans. p. 403.1970. ERNST, A. Exposición Nacional de Venezuela en 1883. Reimpresión. Caracas. Fundación Venezolana para la Salud y la Educación. p. 379.1983. GARBER, E.D. A cytological study of the genus Sorghum Subsection ParaSorghum and EuSorghum. Am. Nat. 78:89-94. 1944. GARBER, E.D. Cytotaxonomy studies in the genus Sorghum. Univ. Calif. Publi. Bot. 23:283-361, 1950. n.v. fide Dogget, H. Sorghum. London. Longmans. p. 403. 1 970. GARBER, E.D. y LA. SNYDER. Cytotaxonomic studies in the genus Sorghum. II. Two new species from Australia. Madroño. 11: 6. 1951. GARBER, E.D. Cytotaxonomic studies in the genus Sorghum. III. The poly. plaid species of the subgenera Parasorghum and Stiposorghum. Bot. Gaz.115336. 1954. GRAY HERBARIUM CARD INDEX. Cambridge Massachusset. HACKEL, E. (Andropogon sorghum. Brot.). In: De Candolle. Monographiae Phanerogamarum 6:500-520.1889. HARLAN, J.R. y J.M.J. DE WET. Toward a rational classification of cultivated plants. Taxon,20:509-517.1971. HARLAN, J.R. Crop et man. American Society of Agronomy. Found for Modem Crop Science Series. Madison. Wisconsin. 295 p. 1975. HOYOS, J. Flora de la Isla de Margarita. Caracas. Soc. y Fund. La Salle. 927 p. 1985. (Monografia No. 34). INDEX KEWENSIS. Plantarum phanerogamarum. 2 vols., 16 suppl. Oxford, 1893-1975. JAKUSHEVSKY, E.S. Varietal composition of sorghum and its use for brceding. Bull. Appl. Genet. Plant Breeding 41: 148-178. n.v. fide Harlan, J.R. Crop et man. Madison. Wisconsin.1975. KARPER. R.E. y A.T. CHRISHOLM. Chromosone numbers in Sorghum. Am. J. of Bot. 23(5):369.374. 1936. KOERNICKE, F. (Andropogon sorghum Brot.), die Mohrhirse n.v. fide Koernicke, F. y H. Werner. Handbuch des Getreidebaues 1 :294-315.1885. KUNTH,C. Nova genera et species plantarum (Andropogon) 1:189-190. 1815. LIANG, G. y J. CASADY. Quantitative presentation on the systematic relationships among twenty-one Sorghum species. Crop Science 6(1):76-79. 1966. LINNAEUS, C. Hortus cliffortianus. Amsteld. (Holcus):468-469.1737. LUCES DE FEBRES, Z. Generos de las gramíneas venezolanas. Bol. Téc. No. 4. Caracas, Ministerio de Agricultura y Cría. p. 141-143. 1942. MALAGUTI, G. Downy mildew disease of Sorghum in the Americas. In: Sorghum diseases. India, ICRISAT. p.150-156.1978. MALAGUTI, G. Los sorgos silvestres como fuente de inóculo del mosaico de la caña de azúcar y del Mildew lanoso en siembras de maíz y sorgo en Venezuela. Phytopathologie Mediterranea 18:205-206. 1979. MALAGUTI, G. The role of collateral hosts in Sorghum Downy Mildew infections in the Americas. Position paper Bellagio Conference. New York. Rockefeller Foundation.18 p. 1980. MURTY, B.R., V. ARUNACHALAM y M.B.L. SAXENA. Classification and catalogue of world collection of Sorghum. Indian J. Genet. 27:1-74. 1967. n.v. fde Dogget, H. Sorghum. London, longmans. p. 403.1970. PIPER, L.V. Andropogon halepense and Andropogon sorghum. Proc. Biol. Soc.Washington.28:25-44.1915. PRIETO, M1. Las melezas del ajonjolí (Sesasum indicum l.) en el Estado Portuguesa. Caracas. Fondo Nacional de Investigaciones Agropecuarias. p. 62-65.1975. ROEMER, JJ. y J.A. SCHULTES. System vegetabilium. Stuttgart. s.e. part 2. p.838.1817. STAPF, O. Sorghum. In: Prain, D. Ed. Flora of tropical Africa. 9:1041S4. 1917. SNOWDEN, J.D. The cultivated race of Sorghum. London. Adlard and son. 274 p.1936. SNOWDEN, J.D. The wild fodder sorghums of section Eusorghum J. Linn. Soc. Lond. Bot.55:191~260.1955. STEUDEL, E.G. Synopsis plantarum glumacearum. Stuttgart. s.e. 1:393. 1854. WILLDENOW, C.L. Caroli a Linné species plantarum. 4 ed. Berlín. s.e. 4 part. 2:931.1805. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||